Essa é uma revisão anterior do documento!
Tabela de conteúdos
Papers
Segregation
Residential segregation in an all-integrationist world
This paper presents a variation of the Schelling [J. Math. Sociol. 1 (1971) 143; T.C. Schelling, Micromotives and Macrobehavior, Norton, New York, 1978] model to show that segregation emerges and persists even if every person in the society prefers to live in a half-black, half-white neighborhood. In contrast to Schelling’s inductive approach, we formulate neighborhood transition as a spatial game played on a lattice graph. The model is rigorously analyzed using techniques recently developed in stochastic evolutionary game theory. We derive our primary results mathematically and use agent-based simulations to explore the dynamics of segregation.
One agent per cell. Simple satisfability function. Schelling: preferences for like-color neighbors at the individual level can be amplified into high levels of segregation.
Language Evolution and Population Dynamics in a System of Two Interacting Species
The evolutionionary origin of inter- and intra-specific cooperation among non-related individuals has been a great challenge for biologists for decades. Recently, the continuous prisoner’s dilemma game has been introduced to study this problem. In function of previous payoffs, individuals can change their cooperative investment iteratively in this model system. Killingback and Doebeli (Am. Nat. 160 (2002) 421–438) have shown analytically that intra-specific cooperation can emerge in this model system from originally non-cooperating individuals living in a non-structured population. However, it is also known froman earlier numerical work that inter-specific cooperation (mutualism) cannot evolve in a very similar model. The only difference here is that cooperation occurs among individuals of different species. Based on the model framework used by Killingback and Doebeli (2002), this Note proves analytically that mutualism indeed cannot emerge in this model system. Since numerical results have revealed that mutualism can evolve in this model system if individuals interact in a spatially structured manner, our work emphasizes indirectly the role of spatial structure of populations in the origin of mutualism.
Each site of the lattice may be empty or it can be occupied by “individuals” of A or by B. One of the four neighbour sites is randomly selected. In the case of empty the individual moves to it. If it is occupied then the two communicate. After the motion there is a probability that reproduction will take place.
Two interacting species which initially speak different languages. The spatial distributions of the species may cause the system to exihbit pattern formation or seggregation. In the most cases the system will arrive at a final state where both languages coexist. The results offer explanation for the existence and origin of synonimous in spoken languages.
Evolutionary Games
On the instability of evolutionary stable strategies in small populations
Gary B. Fogel, Peter C. Andrews, David B. Fogel
Evolutionary stable strategies (ESSs) are often used to explain the behaviors of individuals and species. The analysis of ESSs determines which, if any, combinations of behaviors cannot be invaded by alternative strategies. Two assumptions required to generate an ESS (i.e. an infinite population and payoffs described only on the average) do not hold under natural conditions. Previous experiments have indicated that under more realistic conditions of finite populations and stochastic payoffs, populations may evolve in trajectories that are unrelated to an ESS, even in very simple evolutionary games. The simulations are extended here to small populations with varying levels of selection pressure and mixing levels. The results suggest that ESSs may not provide a good explanation of the behavior of small populations even at relatively low levels of selection pressure and even under persistent mixing. The implications of these results are discussed briefly in light of previous literature which claimed that ESSs generated suitable explanations of real-world data.
{kind=link}
{kind=link}
Mobility and Cooperation: On the Run
In public goods experiments where subjects may change groups, we observe a continual flight of the more cooperative subjects away from the less cooperative ones. The less cooperative subjects attempt to enter cooperative groups in order to free-ride on their contributions.
In the symmetric n-player public good game, each of the n players is endowed with 10 tokens to be allocated between two alternatives, called A and B. Alternative A is private: each token allocated to alternative A yields the player a return of 1. Alternative B is public: each token allocated to alternative B by any of the n players yields each player in the group an individual return of k(n), that depends on the group size n. The individual return per token k(n) is smaller than 1, the return per token in the private alternative A. However, the return per token to the entire group r(n) = nk(n) is greater than 1 (for n > 1). contribution to the private alternative maximizes one’s individual profit but leads to an undesirable outcome for the group.
Each player decides whether he wants to stay with his group, switch to one of the other groups that existed in the previous round, or create a new group.
Experiments with 90 students that know all the rules of the game, but cannot communicate. The major conclusion that we can draw out of these results is that both free-riding and cooperating are active principles.
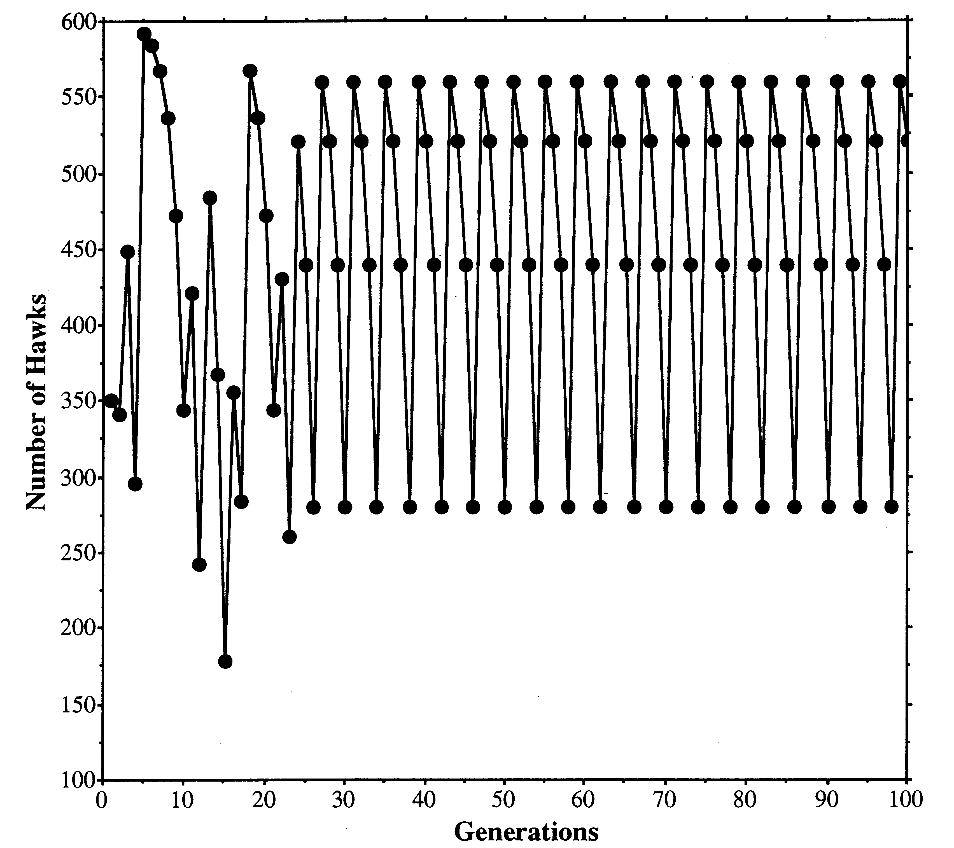
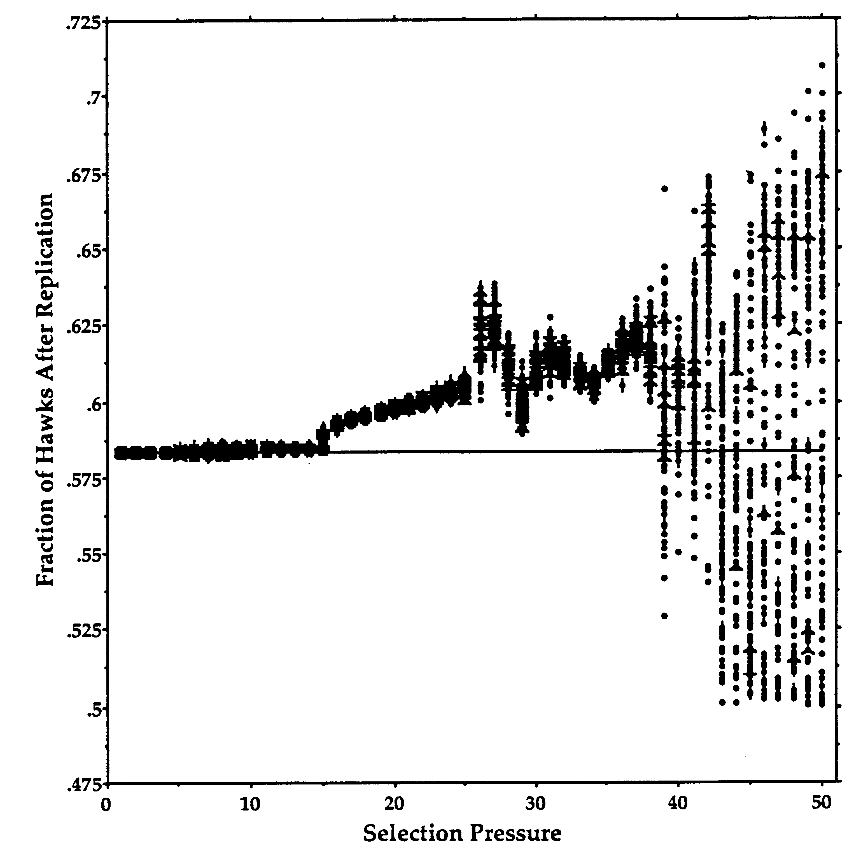
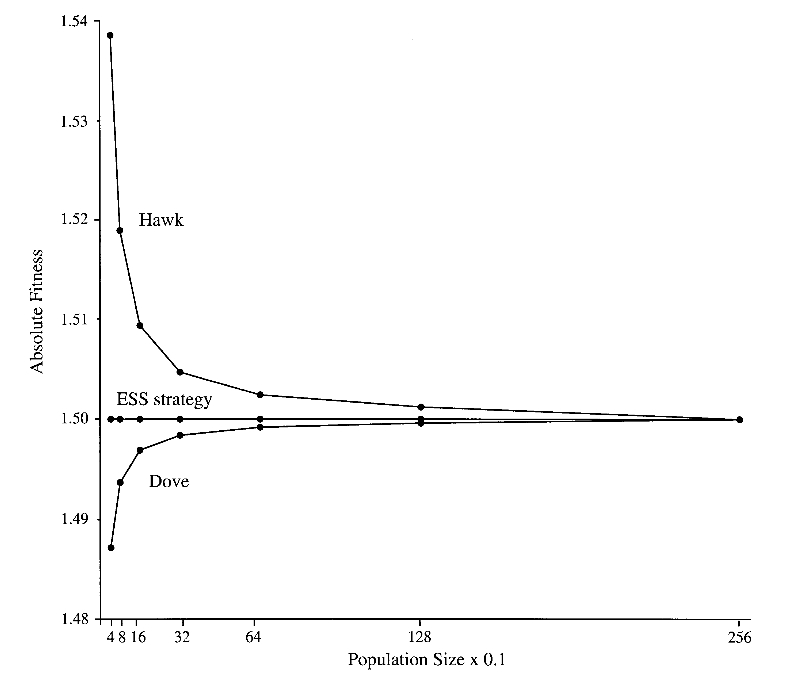
The Evolution of Strategy Variation: Will an ESS Evolve?
Evolutionarily stable strategy (ESS) models are widely viewed as predicting the strategy of an individual that when monomorphic or nearly so prevents a mutant with any other strategy from entering the population. In fact, the prediction of some of these models is ambiguous when the predicted strategy is ‘‘mixed’’, as in the case of a sex ratio, which may be regarded as a mixture of the subtraits ‘‘produce a daughter’’ and ‘‘produce a son.’’ Some models predict only that such a mixture be manifested by the population as a whole, that is, as an ‘‘evolutionarily stable state’’; consequently, strategy monomorphism or polymorphism is consistent with the prediction. The hawk-dove game and the sex-ratio game in a panmictic population are models that make such a ‘‘degenerate’’ prediction. We show here that the incorporation of population finiteness into degenerate models has effects for and against the evolution of a monomorphism (an ESS) that are of equal order in the population size, so that no one effect can be said to predominate. Therefore, we used Monte Carlo simulations to determine the probability that a finite population evolves to an ESS as opposed to a polymorphism. We show that the probability that an ESS will evolve is generally much less than has been reported and that this probability depends on the population size, the type of competition among individuals, and the number of and distribution of strategies in the initial population. We also demonstrate how the strength of natural selection on strategies can increase as population size decreases. This inverse dependency under- scores the incorrectness of Fisher’s and Wright’s assumption that there is just one qualitative relationship between population size and the intensity of natural selection.
{kind=link}
Spatial Games
The Replicator Equation on Graphs
H. Ohtsuki and M. A. Nowak, 2006
We study evolutionary games on graphs. Each player is represented by a vertex of the graph. The edges denote who meets whom. A player can use any one of n strategies. Players obtain a payoff from interaction with all their immediate neighbors. We consider three different update rules, called ‘birth–death’, ‘death–birth’ and ‘imitation’. A fourth update rule, ‘pairwise comparison’, is shown to be equivalent to birth–death updating in our model. We use pair approximation to describe the evolutionary game dynamics on regular graphs of degree k. In the limit of weak selection, we can derive a differential equation which describes how the average frequency of each strategy on the graph changes over time. Remarkably, this equation is a replicator equation with a transformed payoff matrix. Therefore, moving a game from a well-mixed population (the complete graph) onto a regular graph simply results in a transformation of the payoff matrix. The new payoff matrix is the sum of the original payoff matrix plus another matrix, which describes the local competition of strategies. We discuss the application of our theory to four particular examples, the Prisoner’s Dilemma, the Snow-Drift game, a coordination game and the Rock–Scissors–Paper game.
Bij (the transformation in the payoff matrix) can be calculated because there is a fixed neighbourhood size for all the graph. This work generalizes some works presented in the literature, including Hauert and Doebeli Spatial structure often inhibits the evolution of cooperation in the snowrift game (Nature)
The evolution of interspecific mutualisms
M. Doebeli and N. Knowlton, 1998
Interspecific mutualisms are widespread, but how they evolve is not clear. The Iterated Prisoner’s Dilemma is the main theoretical tool to study cooperation, but this model ignores ecological differences between partners and assumes that amounts exchanged cannot themselves evolve. A more realistic model incorporating these features shows that strategies that succeed with fixed exchanges (e.g., Tit-for-Tat) cannot explain mutualism when exchanges vary because the amount exchanged evolves to 0. For mutualism to evolve, increased investments in a partner must yield increased returns, and spatial structure in competitive interactions is required. Under these biologically plausible assumptions, mutualism evolves with surprising ease. This suggests that, contrary to the basic premise of past theoretical analyses, overcoming a potential host’s initial defenses may be a bigger obstacle for mutualism than the subsequent recurrence and spread of noncooperative mutants.
{kind=link}
Spatial Mendelian Games
J. Radcliffe and L. Rass, 1998
This paper considers complex models arising in sociobiology. These combine genetic and strategic aspects to model the effect of gene-linked strategies on the ability of individuals to survive to maturity, mate and produce offspring. Several important models considered in the literature are generalised and extended to incorporate a spatial aspect. Individuals are allowed to migrate. Contests, e.g. for food or amongst males for females, take place locally. The choice of the point at which the population structure is measured affects the complexity of the equations describing the system, although it is possible to utilise any point in the life cycle. For our spatial models the simplest approach is to measure the population structure immediately after migration. A saddle point method, developed by the authors, has previously been used to obtain results for simple discrete time spatial models. It is utilised here to obtain the speed of first spread of a new gene-linked strategy for the much more complex sociobiological models included in this paper. This demonstrates the wide-ranging applicability and power of the method.
Games against vicinity. The game has two pure strategies s1 and s2. Population: A1A1 (1), A1A2 (2), A2A2 (3). (1) plays s1, (2) plays s2 and (3) plays s1 with probability p and s2 with 1-p. Reaping occours to reduce the population to the carrying capacity of the habitat. A new population is generated and the previous one is removed. There is a probability density function for migration.
Evolution of Cooperation in Spatially Structured Populations
K. Brauchli and T. Killingback and M. Doebeli, 1999
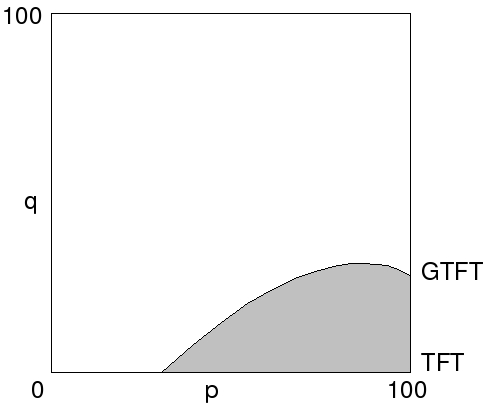
Using a spatial lattice model of the Iterated Prisoner's Dilemma we studied the evolution of cooperation within the strategy space of all stochastic strategies with a memory of one round. Comparing the spatial model with a randomly mixed model showed that (1) there is more cooperative behaviour in a spatially structured population, (2) PAVLOV and generous variants of it are very successful strategies in the spatial context and (3) in spatially structured populations evolution is much less chaotic than in unstructured populations. In spatially structured populations, generous variants of PAVLOV are found to be very successful strategies in playing the Iterated Prisoner's Dilemma. The main weakness of PAVLOV is that it is exploitable by defective strategies. In a spatial context this disadvantage is much less important than the good error correction of PAVLOV, and especially of generous PAVLOV, because in a spatially structured population successful strategies always build clusters.
pavlov strategy means: “win stay loose shift”. in a Generous strategy there is a minor probability of not defecting even when the strategy forces it. GTFT
{kind=link}
The Spatial Ultimatum Game
In the ultimatum game, two players are asked to split a certain sum of money. The proposer has to make an offer. If the responder accepts the offer, the money will be shared accordingly. If the responder rejects the offer, both players receive nothing. The rational solution is for the proposer to offer the smallest possible share, and for the responder to accept it. Human players, in contrast, usually prefer fair splits. In this paper, we use evolutionary game theory to analyse the ultimatum game. We first show that in a nonspatial setting, natural selection chooses the unfair, rational solution. In a spatial setting, however, much fairer outcomes evolve.
Players arranged on a two-dimensional square lattice. Each player interacts with his neighbours. Experiments on the UG shed a striking light on our mental equipment for social and economic life. Who do fairness considerations matter more, to many of us, than rational utility maximization? Spatial population structure can have important effects on the evolutionary outcome of the ultimatum game.
Disordered environments in spatial games
M. H. Vainstein and J. J. Arenzon, 2001
The Prisoner’s dilemma is the main game theoretical framework in which the onset and maintainance of cooperation in biological populations is studied. In the spatial version of the model, we study the robustness of cooperation in heterogeneous ecosystems in spatial evolutionary games by considering site diluted lattices. The main result is that, due to disorder, the fraction of cooperators in the population is enhanced. Moreover, the system presents a dynamical transition at P, separating a region with spatial chaos from one with localized, stable groups of cooperators.
We allow that some of the sites may be empty. No empty site will be ever filled. In the simulations, averages are taken from 100 samples.
{kind=link}
Spatial Evolutionary Games of Interaction among Generic Cancer Cells
L.A. Bach and D. J. T. Sumpter and J. Alsner and V. Loeschke, 2003
Evolutionary game models of cellular interactions have shown that heterogeneity in the cellular genotypic composition is maintained through evolution to stable coexistence of growth-promoting and non-promoting cell types. We generalise these mean-field models and relax the assumption of perfect mixing of cells by instead implementing an individual-based model that includes the stochastic and spatial effects likely to occur in tumours. The scope for coexistence of genotypic strategies changed with the inclusion of explicit space and stochasticity. The spatial models show some interesting deviations from their mean-field counterparts, for example the possibility of altruistic (paracrine) cell strategies to thrive. Such effects can however, be highly sensitive to model implementation and the more realistic models with semi-synchronous and stochastic updating do not show evolution of altruism. We do find some important and consistent differences between the spatial and mean-field models, in particular that the parameter regime for coexistence of growth-promoting and nonpromoting cell types is narrowed. For certain parameters in the model a selective collapse of a generic growth promoter occurs, hence the evolutionary dynamics mimics observable in vivo tumour phenomena such as (therapy induced) relapse behaviour. Our modelling approach differs from many of those previously applied in understanding growth of cancerous tumours in that it attempts to account for natural selection at a cellular level. This study thus points a new direction towards more plausible spatial tumour modelling and the understanding of cancerous growth.
Finding a Nash Equilibrium in Spatial Games is an NP-Complete Problem
R. Baron, J. Durieu, H. Haller and P. Solal, 2004
we consider the class of (finite) spatial games. We show that the problem of determining wether there is a Nash Equilibrium in which each player has a payoff of at leas k is NP-Complete as a function of the number of players. When each player has two strategies and the base game is an anti-coordination game, the problem is decidable in polynomial time.
Prisoner’s dilemma on dynamic networks under perfect rationality
C. Biely and K. Dragosits and S. Thurner, 2005
We consider the prisoner’s dilemma being played repeatedly on a dynamic network, where agents may choose their actions as well as their co-players. In the course of the evolution of the system, agents act fully rationally and base their decisions only on local information. Individual decisions are made such that links to defecting agents are resolved and that cooperating agents build up links, as new interrelations are established via a process of recommendation. The dynamics introduced thereby leads to periods of growing cooperation and growing total linkage, as well as to periods of increasing defection and decreasing total linkage, quickly following each other if the players are perfectly synchronized. The cyclical behavior is lost and the system is stabilized when agents react ’slower’ to new information. Our results show, that within a fully rational setting in a licentious society, the prisoner’s dilemma leads to overall cooperation and thus loses much of its fatality when a larger range of dynamics of social interaction is taken into account. We also comment on the emergent network structures.
{kind=link}
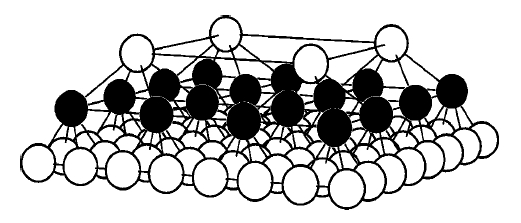
Evolutionary prisoner’s dilemma game on hierarchical lattices
An evolutionary prisoner’s dilemma sPDd game is studied with players located on a hierarchical structure of layered square lattices. The players can follow two strategies fD sdefectord and C scooperatordg and their income comes from PD games with the “neighbors.” The adoption of one of the neighboring strategies is allowed with a probability dependent on the payoff difference. Monte Carlo simulations are performed to study how the measure of cooperation is affected by the number of hierarchical levels sQd and by the temptation to defect. According to the simulations the highest frequency of cooperation can be observed at the top level if the number of hierarchical levels is low sQ,4d. For larger Q, however, the highest frequency of cooperators occurs in the middle layers. The four-level hierarchical structure provides the highest average stotald income for the whole community.
{kind=link}
Evolutionary Prisioner's Dilemma in Random Graphs
We study an evolutionary version of the spatial prisoner’s dilemma game (SPD), where the agents are placed in a random graph. For graphs with fixed connectivity, alpha, we show that for low values of alpha the final density of cooperating agents, Pc depends on the initial conditions. However, if the graphs have large connectivities Pc is independent of the initial conditions. We characterize the phase diagram of the system, using both, extensive numerical simulations and analytical computations. It is shown that two well defined behaviors are present: a Nash equilibrium, where the final density of cooperating agents Pc is constant, and a non-stationary region, where Pc fluctuates in time. Moreover, we study the SPD in Poisson random graphs and find that the phase diagram previously developed looses its meaning. In fact, only one regime may be defined. This regime is characterized by a non stationary final state where the density of cooperating agents varies in time.
Search for models able to account for the complex behaviour in many biological, economical and social systema has lead to an intense research activity in the last years.
Players on nodes playing Prisioner's dilemma against their neighbours. Random Graph 1: Bethe Lattice, sites with fixed and equal number alpha of neighbors. RG 2: Poisson random graph, links distributed with a mean value alpha.
The evolution of cooperation depends on the connectivity and on the game payoff, but it is independent of the initial conditions. Poisson random graphs lead to more cooperation then with graphs of fixed degree.
The Iterated Continuous Prisioner's Dilemma Game Cannot Explain the Evolution of Interspecific Mutualism in Unstructured Populations
The evolutionionary origin of inter- and intra-specific cooperation among non-related individuals has been a great challenge for biologists for decades. Recently, the continuous prisoner’s dilemma game has been introduced to study this problem. In function of previous payoffs, individuals can change their cooperative investment iteratively in this model system. Killingback and Doebeli (Am. Nat. 160 (2002) 421–438) have shown analytically that intra-specific cooperation can emerge in this model system from originally non-cooperating individuals living in a non-structured population. However, it is also known froman earlier numerical work that inter-specific cooperation (mutualism) cannot evolve in a very similar model. The only difference here is that cooperation occurs among individuals of different species. Based on the model framework used by Killingback and Doebeli (2002), this Note proves analytically that mutualism indeed cannot emerge in this model system. Since numerical results have revealed that mutualism can evolve in this model system if individuals interact in a spatially structured manner, our work emphasizes indirectly the role of spatial structure of populations in the origin of mutualism.
It is highly improbable that mutualistic interaction would emerge in this model system. We emphasize here that our analysis is restricted only to the large population limit […], which could not exclude the evolution of mutualism in small populations.
On the other side, this letter emphasizes indirectly the important role of spatial structures in the evolution of mutualism.
Evolutionary Game Theory in an Agent-Based Brain Tumor Model: Exploring the 'Genotype-Fenotype' Link
To investigate the genotype–phenotype link in a polyclonal cancer cell population, here we introduce evolutionary game theory into our previously developed agent-based brain tumor model. We model the heterogeneous cell population as a mixture of two distinct genotypes: the more proliferative Type A and the more migratory Type B. Our agent-based simulations reveal a phase transition in the tumor’s velocity of spatial expansion linking the tumor fitness to genotypic composition. Specifically, velocity initially falls as rising payoffs reward the interactions among the more stationary Type A cells, but unexpectedly accelerates again when these A–A payoffs increase even further. At this latter accelerating stage, fewer migratory Type B cells appear to confer a competitive advantage in terms of the tumor’s spatial aggression over the overall numerically dominating Type A cells, which in turn leads to an acceleration of the overall tumor dynamics while its surface roughness declines. We discuss potential implications of our findings for cancer research.
Heterogenous cell population as a mixture of two distinct genotypes: the more proliferative (A) and the more migratory (B)
Cells can perform one of these actions: proliferate, invade, or turn quiescent. Both proliferation and migration occur within the same time scale. Cells that do not proliferate or migrate automatically enter a reversible, quiescent state.
At any given time, a lattice site can be either empty or occupied by at most one single tumor cell.
There are two nutrient sources (representing e.g. cerebral blood vessels) at the grid center and in the middle of the mortheast quadrant.
Cells that do not proliferate or migrate automatically enter a reversible, quiescent state.
Spatial Effects in Social Dilemmas
Social dilemmas and the evolutionary conundrum of cooperation are traditionally studied through various kinds of game theoretical models such as the prisoner’s dilemma, public goods games, snowdrift games or by-product mutualism. All of them exemplify situations which are characterized by different degrees of conflicting interests between the individuals and the community. In groups of interacting individuals, cooperators produce a common good benefitting the entire group at some cost to themselves, whereas defectors attempt to exploit the resource by avoiding the costly contributions. Based on synergistic or discounted accumulation of cooperative benefits a unifying theoretical framework was recently introduced that encompasses all games that have traditionally been studied separately (Hauert, Michor, Nowak, Doebeli, 2005. Synergy and discounting of cooperation in social dilemmas. J. Theor. Biol., in press.). Within this framework we investigate the effects of spatial structure with limited local interactions on the evolutionary fate of cooperators and defectors. The quantitative effects of space turn out to be quite sensitive to the underlying microscopic update mechanisms but, more general, we demonstrate that in prisoner’s dilemma type interactions spatial structure benefits cooperation—although the parameter range is quite limited—whereas in snowdrift type interactions spatial structure may be beneficial too, but often turns out to be detrimental to cooperation.
(M. Smith 95) All major transitions in evolution can be reduced to successfull resolutions of social dilemmas under Darwinian selection
(Moran 62) process: a focal individual is randomly chosen for reproduction with a probability proportional to its fitness. Another randomly chosen is eliminated and replaced by an offspring of the focal individual. perhaps also choose based on fitness?
(Otsuki 05) process: like Moran but assuming a death-birth instead of a birth-death process.
Spatial structure enables cooperators to thrive by forming clusters and thereby reducing exploitation by defectors.
Each additional benefit may be discounted or synergistically enhanced by a factor w. [Forçando a barra para a cooperação?  : Verificar no artigo]
: Verificar no artigo]
The geometrical patterns of cooperation evolution in the spatial prisoner’s dilemma: An intra-group model
R. O. S. Soares and A. S. Martine, 2006
The prisoner’s dilemma (PD) deals with the behavior conflict between two agents, who can either cooperate (cooperators) or defect. If both agents cooperate (defect), they have a unitary (null) payoff. Otherwise the payoff is T for the defector and null for the cooperator. The temptation T to defect is the only free parameter in the model. Here the agents are represented by the cells of a LxL lattice. The agent behaviors are initially randomly distributed according to an initial proportion of cooperators Pc(0). Each agent has no memory of previous behaviors and plays the PD with his/her eight nearest neighbors. At each generation, the considered agent copies the behavior of those who have secured the highest payoff. Once the PD conflict has been established (1<T<2), this system shows that cooperation among agents may emerge even for reasonably high T values giving rise to the well-known strategy: join to conquer, fight to share. Contrary to previous studies, in which the lattice cells are viewed as groups and are allowed to self-interact (inter-group situation), here the cells are viewed as individuals and are not allowed to self-interact (intra-group situation). Although the short time and asymptotic behavior of Pc are similar in both cases, the intermediate behavior is different. Oscillations in the intra-group Pc(t) forbids data collapse. The cooperators clusters geometrical configurations are distinct between inter and intra-group models, which explains the Pc(t) differences.
Evolutionary prisoner’s dilemma game with dynamic preferential selection
Z. Wu, X. Xu, S. Wang and Y. Wang, 2006
A modified prisoner’s dilemma game is numerically investigated on disordered square lattices characterized by a fi portion of random rewired links with four fixed number of neighbors of each site. The players interacting with their neighbors can either cooperate or defect and update their states by choosing one of the neighboring and adopting its strategy with a probability depending on the payoff difference. The selection of the neighbor obeys a dynamic preferential rule: the more frequency a neighbor’s strategy was adopted in the previous rounds, the larger probability it was picked. It is found that this simple rule can promote greatly the cooperation of the whole population with disordered spatial distribution. Dynamic preferential selection are necessary to describe evolution of a society whose actions may be affected by the results of former actions of the individuals in the society. Thus introducing such selection rule helps to model dynamic aspects of societies.
Prisoner's Dilemma Game with Heterogeneous Influential Effect on Regular Small-World Networks
Wu Zhi-Xi and Xu Xin-Jian and Wang Ying-Hai, 2006
The effect of heterogeneous infuence of different individuals on the maintenance of co-operative behaviour is studied in an evolutionary Prisoner's Dilemma game with players located on the sites of regular small-world networks. The players interacting with their neighbours can either co-operate or defect and update their states by choosing one of the neighbours and adopting its strategy with a probability depending on the payoff difference. The selection of the neighbour obeys a preferential rule: the more influential a neighbour, the larger the probability it is picked. It is found that this simple preferential selection rule can promote continuously the co-operation of the whole population with the strengthening of the disorder of the underlying network.
{kind=link}
Agents
Defining Spatial Agents
The concept of agent is becoming increasingly important not only in research (where it has been in use for some time) but also now in commercial applications. However, an agent may represent many d ifferent things according to the people that implement and use them. Based on the concept of agent we define Spatial Agents as agents that make spatial concepts computable. By implementing spatial agents we hope to solve the following problems : Locating and retrieving Spatial Information in large networks (and specifically the Internet), Facilitate the handling of a GIS user interface, Implementing improved spatial tasks and Creating interfaces between GIS and specific software packages. We discuss what are the necessary qualities that a development tool should have to qualify for agent development. We select some of the most prominent tools currently used and try to choose which are best suited for the development of spatial agents. Finally, we reflect on the design of spatial agents that will solve the problems mentioned above and present a prototype of an Interface Agent for the Drawing tool of the Smallworld GIS.
Out-of-Equilibrium Economics and Agent-Based Modeling
Standard neoclassical economics asks what agents’ actions, strategies, or expectations are in equilibrium with (consistent with) the outcome or pattern these behaviors aggregatively create. Agent-based computational economics enables us to ask a wider question: how agents’ actions, strategies, or expectations might react to—might endogenously change with—the patterns they create. In other words, it enables us to examine how the economy behaves out of equilibrium, when it is not at a steady state. This out-of-equilibrium approach is not a minor adjunct to standard economic theory; it is economics done in a more general way. When examined out of equilibrium, economic patterns sometimes simplify into a simple, homogeneous equilibrium of standard economics; but just as often they show perpetually novel and complex behavior. The static equilibrium approach suffers two characteristic indeterminacies: it cannot easily resolve among multiple equilibria; nor can it easily model individuals’ choices of expectations. Both problems are ones of formation (of an equilibrium and of an “ecology” of expectations, respectively), and when analyzed in formation—that is, out of equilibrium—these anomalies disappear.
Modelling adaptive, spatially aware, and mobile agents: Elk migration in Yellowstone
D. A. Bennett and W. Tang, 2006
The potential utility of agent-based models of adaptive, spatially aware, and mobile entities in geographic and ecological research is considerable. Developing this potential, however, presents significant challenges to geographic information science. Modelling the spatio-temporal behaviour of individuals requires new representational forms that capture how organisms store and use spatial information. New procedures must be developed that simulate how individuals produce bounded knowledge of geographical space through experiential learning, adapt this knowledge to continually changing environments, and apply it to spatial decision-making processes. In this paper, we present a framework for the representation of adaptive, spatially aware, and mobile agents. To provide context to this research, a multiagent model is constructed to simulate the migratory behaviour of elk (Cervus elaphus) on Yellowstone’s northern range. In this simulated environment, intelligent agents learn in ways that enable them to mimic real-world behaviours and adapt to changing landscapes.
In mechanistic models, it is assumed that agent's spatial knowledge is limited to its perceptual range and that it has limited ability to remember past experiences or predict future states. two forms of spatial memory: episodic and reference.
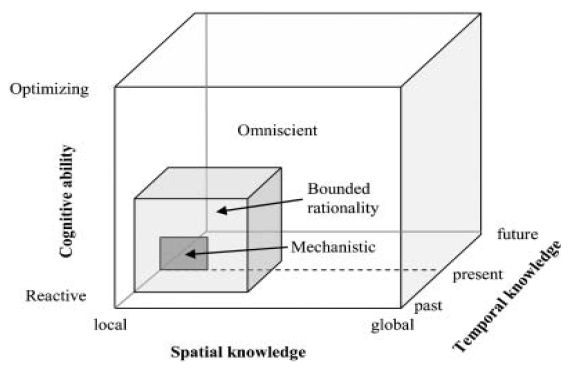{kind=link}
Spatial Behavior in Groups: an Agent-Based Approach
F. S. Beltran, L. Salas and V. Quera, 2006
We present an agent-based model with the aim of studying how macro-level dynamics of spatial distances among interacting individuals in a closed space emerge from micro-level dyadic and local interactions. Our agents moved on a lattice (referred to as a room) using a model implemented in a computer program called P-Space in order to minimize their dissatisfaction, defined as a function of the discrepancy between the real distance and the ideal, or desired, distance between agents. Ideal distances evolved in accordance with the agent's personal and social space, which changed throughout the dynamics of the interactions among the agents. In the first set of simulations we studied the effects of the parameters of the function that generated ideal distances, and in a second set we explored how group macro-level behavior depended on model parameters and other variables. We learned that certain parameter values yielded consistent patterns in the agents' personal and social spaces, which in turn led to avoidance and approaching behaviors in the agents. We also found that the spatial behavior of the group of agents as a whole was influenced by the values of the model parameters, as well as by other variables such as the number of agents. Our work demonstrates that the bottom-up approach is a useful way of explaining macro-level spatial behavior. The proposed model is also shown to be a powerful tool for simulating the spatial behavior of groups of interacting individuals.
it cites the example of a party, where players move forming groups. how to describe the behaviour?
move to empty cells on a lattice in accordance with an established rule: each agent could move to a cell in its Moore neighborhood (defined as a 3-cell by 3-cell square with the agent's current location in the center) where the sum of attitude values was maximized.
LUCC
Spatially explicit experiments for the exploration of land-use decision-making dynamics
T. P. Evans and W. Sun and H. Kelley
We explore the special outcomes of decision-making through two laboratorybased experiments, one with a homogenous land suitability surface and another with a heterogeneous suitability surface. Subjects make resource allocation decisions on an abstract landscape and are given a monetary incentive to maximize their revenue during the experiment. These experimental results are compared with simulation output from an agent-based model run on the same abstract landscape that uses a utility-maximizing agent. The main findings are: (1) landscapes produced by subjects result in greater patchiness and more edge than the utility-maximization agent predicts; (2) there is considerable diversity in the decisions subjects make despite the relatively simple decision-making context; and (3) there is greater deviation of subject revenue from the maximum potential revenue in early rounds of the experiment compared with later rounds, demonstrating the challenge of making optimal decisions with little historical context. The findings demonstrate the value of using non-maximizing agents in agent-based models of land-cover change and the importance of acknowledging actor heterogeneity in land-change systems.
We use the term 'experiment' to refer to a laboratory experiment where human subjects are faced with a specific decision-making task rather than to refer to a model run or simulation.
Game theoretic approaches present simplified decision-making settings that are then, with caution, extrapolated to real-world contexts.
They cite some works of games, but they are 'RPG', instead of 'experiments'.
Others
A random matching theory
C.D. Aliprantis and G. Camera and D. Puzzellob, 2006
We develop theoretical underpinnings of pairwise random matching processes. We formalize the mechanics of matching, and study the links between properties of the different processes and trade frictions. A particular emphasis is placed on providing a mapping between matching technologies and informational constraints.
The technical procedure that we use to construct any random matching process involves three basic steps. The first step is to specify how to divide the population in each period into spatially separated clusters of agents. To do so, we use partitional correspondences. Then, one must define and calculate all possible ways to form pairs in each cluster. In this case, we resort to using a class of permutation functions, the so-called involutions. Finally, for each period one must specify a probability measure over all possible pairings, for each cluster.
Provides a formal definition for spatial clusters. Definition of bilateral matching.
(the population has infinitely many agents) That is, not only every pair meets again with probability zero (which we called weak anonymity), but also we show how to ensure that every pair does not share past partners, etc. In brief, we construct matches in which agents are completely isolated from an informational standpoint.
Hybrid cellular automata model for Railway Transportation System and its implementation on GIs
Qin Yong Wang YingJie Jia Li-Min
They present an implementation, with little details, and show the traffic in China railways along common days and on holidays. The paper quality is bad, and I cannot see the details of the automata.
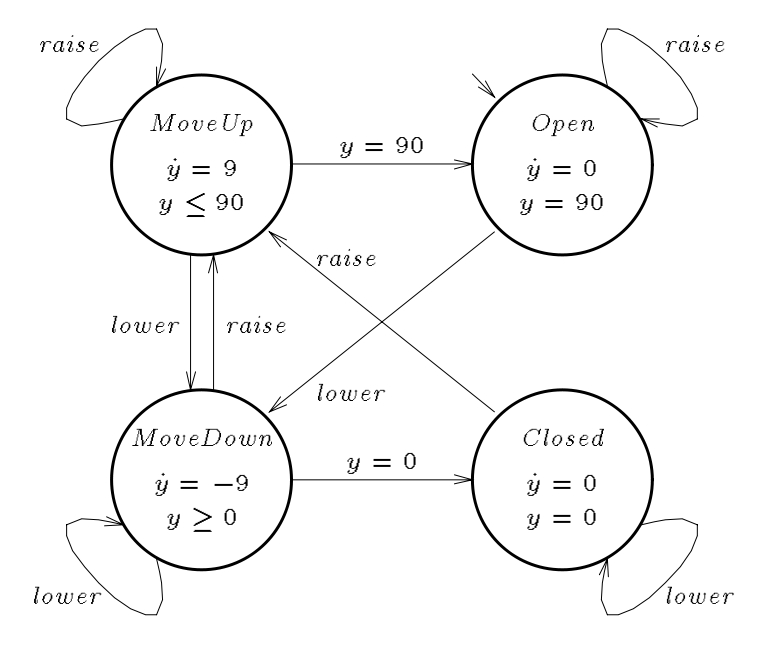
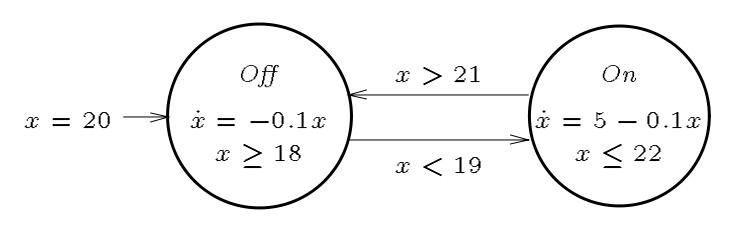
The Theory of Hybrid Automata
{kind=link}
{kind=link}
A paradigmatic example of a mixed discrete-continuous system is a digital con- troller of an analog plant. The discrete state of the controller is modeled by the vertices of a graph (control modes), and the discrete dynamics of the controller is modeled by the edges of the graph (control switches). The continuous state of the plant is modeled by points in Rn, and the continuous dynamics of the plant is modeled by ow conditions such as di erential equations. The behavior of the plant depends on the state of the controller: each control mode determines a flow condition, and each control switch may cause a discrete change in the state of the plant, as determined by a jump condition. Dually, the behavior of the controller depends on the state of the plant: each control mode continuously observes an invariant condition of the plant state, and by violating the invariant condition, a continuous change in the plant state will cause a control switch.
A hybrid automaton H consists of the following components: variables, control graph (control modes=V, control switches=E), initial, invariant, and flow conditions, jump conditions (one for each control switch), and events.